

Автор: Нетрусов А.И.Статей: 2Бактерии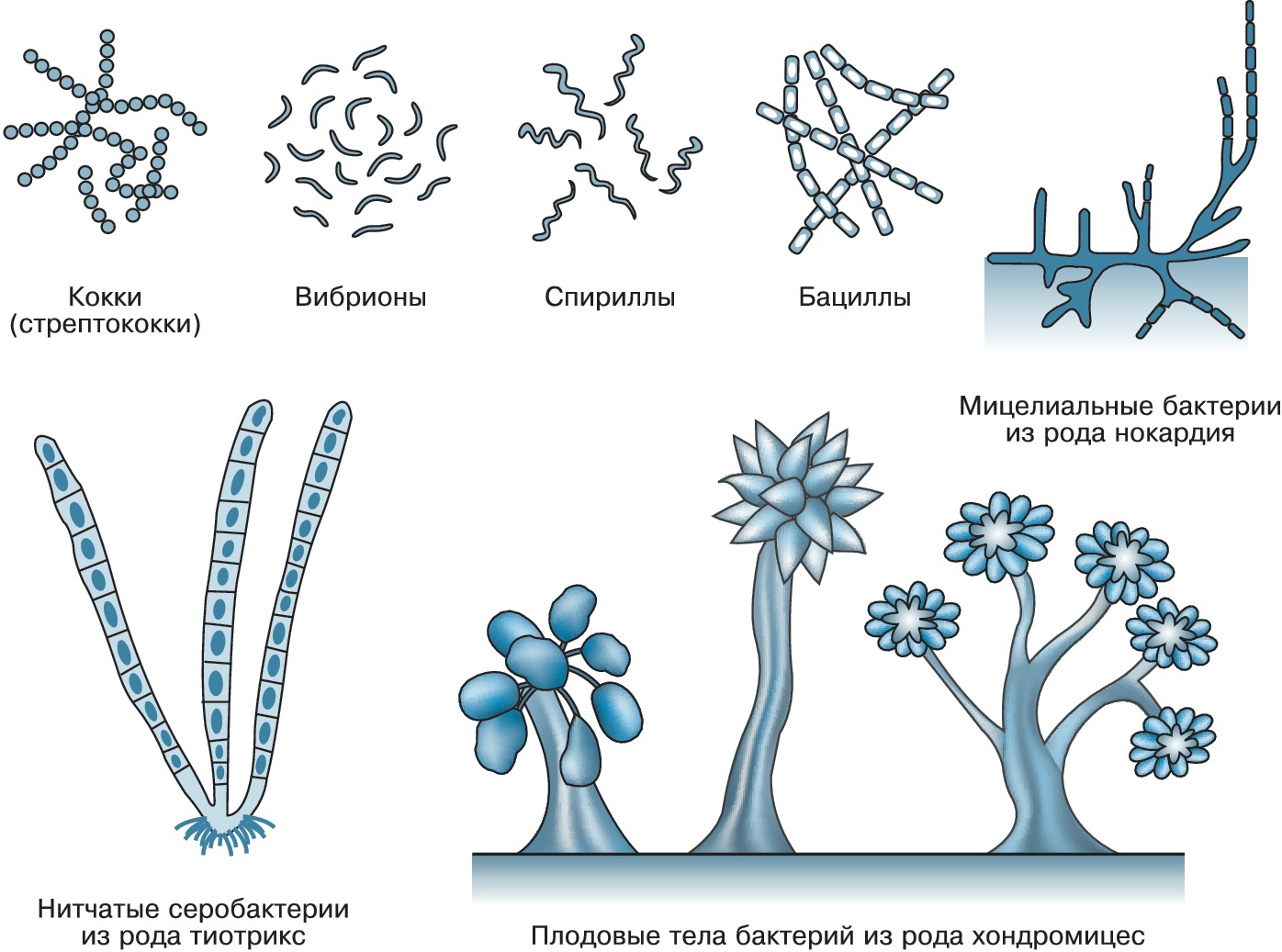 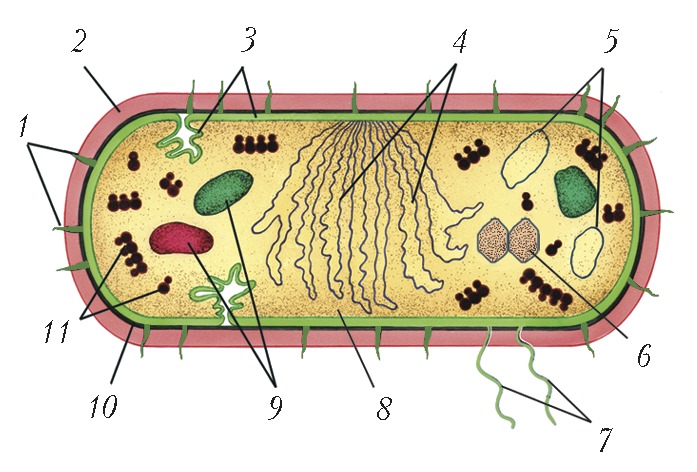 Основные формы бактерий БАКТЕРИИАвторы: А.И. НетрусовБАКТЕРИИ (от греч. βαϰτ ή ριον - палочка) (эубактерии, настоящие, или истинные, бактерии), микроорганизмы с прокариотным типом строения клетки: генетич. аппарат у них не заключён в обособленное мембраной клеточное ядро. Размеры и формы клетокБольшинство Б. - одноклеточные организмы размером 0,2-10,0 мкм. Встречаются среди Б. и «карлики», т. н. нанобактерии (ок. 0,05 мкм), и «гиганты», напр. Б. родов Achromatium и Macromonas (длина до 100 мкм), обитатель кишечника рыбы-хирурга Epulopiscium fishelsoni (длина до 600 мкм) и выделенная из прибрежных морских вод Намибии и Чили Thiomargarita namibiensis (до 800 мкм). Чаще бактериальная клетка имеет вид палочки, сферическую (кокки) или извитую (вибрионы, спириллы и спирохеты) форму. Обнаружены виды с треугольными, квадратными, звездчатыми и плоскими (тарелкообразными) клетками. Некоторые Б. содержат цитоплазматич. выросты - простеки. Б. могут быть одиночными, образовывать пары, короткие и длинные цепочки, грозди, формировать пакеты по 4, 8 и более клеток (сарцины), розетки, сети и мицелий (актиномицеты). Известны также многоклеточные формы, образующие прямые и ветвящиеся трихомы (микроколонии). Встречаются как подвижные, так и неподвижные Б. Первые чаще всего перемещаются с помощью жгутиков, иногда путём скольжения клеток (миксобактерии, цианобактерии, спирохеты и др.). Известно также «прыгающее» движение, природа которого не выяснена. Для подвижных форм описаны явления активного движения в ответ на действия физических или химических факторов.Химический состав и строение клетокБактериальная клетка обычно на 70-80% состоит из воды. В сухом остатке на долю белка приходится 50%, компонентов клеточной стенки 10-20%, РНК 10-20%, ДНК 3- 4% и липидов 10%. При этом в ср. количество углерода составляет 50%, кислорода 20%, азота 14%, водорода 8%, фосфора 3%, серы и калия по 1%, кальция и магния по 0,5% и железа 0,2%. За немногими исключениями (микоплазмы) клетки Б. окружены клеточной стенкой, которая определяет форму Б. и выполняет механич. и важные физиологич. функции. Осн. её компонентом является сложный биополимер муреин (пептидогликан). В зависимости от особенностей состава и строения клеточной стенки Б. по-разному ведут себя при окрашивании по методу Х.К. Грама (дат. учёного, предложившего способ окраски), что послужило основанием для деления Б. на грамположительные, грамотрицательные и на лишённые клеточной стенки (напр., микоплазмы). Первые отличаются большим (до 40 раз) содержанием муреина и толстой стенкой; у грамотрицательных она существенно тоньше и покрыта снаружи внешней мембраной, состоящей из белков, фосфолипидов и липополисахаридов и, по-видимому, участвующей в транспорте веществ. У мн. Б. на поверхности имеются ворсинки (фимбрии, пили) и жгутики, обеспечивающие их движение. Часто клеточные стенки Б. окружены слизистыми капсулами разл. толщины, образованными гл. обр. полисахаридами (иногда гликопротеинами или полипептидами). У ряда Б. обнаружены также т. н. S-слои (от англ. surface - поверхность), выстилающие наружную поверхность клеточной оболочки равномерно упакованными белковыми структурами правильной формы. Цитоплазматич. мембрана, отделяющая цитоплазму от клеточной стенки, служит осмотическим барьером клетки, регулирует транспорт веществ; в ней осуществляются процессы дыхания, азотфиксации, хемосинтез и др. Нередко она образует впячивания - мезосомы. С цитоплазматич. мембраной и её производными связан также биосинтез клеточной стенки, спорообразование и т. д. К ней прикреплены жгутики, геномная ДНК. Схема строения бактериальной клетки: 1 - пили; 2 - слизистая капсула; 3 - цитоплазматическая мембрана; 4 - геномная ДНК (нуклеоид); 5 - плазмиды; 6 - карбоксисомы;... Бактериальная клетка организована довольно просто. В цитоплазме многих Б. имеются включения, представленные разл. рода пузырьками (везикулами), образованными в результате впячивания цитоплазматич. мембраны. Для фототрофных, нитрифицирующих и метанокисляющих Б. характерна развитая сеть цитоплазматич. мембран в виде неразделённых пузырьков, напоминающих граны хлоропластов эукариот. В клетках некоторых обитающих в воде Б. имеются газовые вакуоли (аэросомы), выполняющие роль регуляторов плотности; у мн. Б. обнаружены включения запасных веществ - полисахаридов, поли- β -гидроксибутирата, полифосфатов, серы и др. В цитоплазме присутствуют также рибосомы (от 5 до 50 тыс.). У некоторых Б. (напр., у мн. цианобактерий) имеются карбоксисомы - тельца, в которые заключён фермент, участвующий в фиксации CO 2. В т. н. параспоральных тельцах некоторых спорообразующих Б. содержится токсин, убивающий личинок насекомых. Геном Б. (нуклеоид) представлен кольцевой молекулой ДНК, которую часто называют бактериальной хромосомой. Для бактериального генома характерно объединение многих функционально связанных генов в т. н. опероны. Кроме того, в клетке могут присутствовать внехромосомные генетич. элементы - ДНК плазмид, которые несут неск. полезных для Б. генов (в т. ч. гены устойчивости к антибиотикам). Она может существовать автономно или временно включаться в хромосому. Но иногда, в результате мутаций, эта ДНК теряет способность выходить из хромосомы и становится постоянным компонентом генома. Появление новых генов может быть также обусловлено генетич. переносом в результате однонаправленной передачи ДНК из клетки-донора в клетку-реципиент (аналог полового процесса). Такая передача может осуществляться при прямом контакте двух клеток (конъюгация), при участии бактериофагов (трансдукция) или путём попадания генов в клетку из внешней среды без межклеточного контакта. Всё это имеет большое значение для микроэволюции Б. и приобретения ими новых свойств.РазмножениеБольшинство Б. размножаются путём деления надвое, реже почкованием, а некоторые (напр., актиномицеты) - с помощью экзоспор или обрывков мицелия. Известен способ множественного деления (с образованием мелких репродуктивных клеток-баеоцитов у ряда цианобактерий). Многоклеточные прокариоты могут размножаться отделением от трихом одной или нескольких клеток. Некоторые Б. характеризуются сложным циклом развития, в процессе которого могут меняться морфология клеток и образовываться покоящиеся формы: цисты, эндоспоры, акинеты. Миксобактерии способны образовывать плодовые тела, часто причудливых конфигураций и окрасок. Отличит. особенностью Б. является способность к быстрому размножению. Напр., время удвоения клеток кишечной палочки (Escherichia coli) составляет 20 мин. Подсчитано, что потомство одной клетки в случае неограниченного роста уже через 48 ч превысило бы массу Земли в 150 раз.Условия обитанияБ. приспособились к разным условиям существования. Они могут развиваться в диапазоне темп-р от -5 (и ниже) до 113 °С. Среди них выделяют: психрофилов, растущих при темп-ре ниже 20 °С (для Bacillus psichrophilus, напр., предельная темп- ра роста -10 °С), мезофилов (оптимум роста при 20-40°С), термофилов (50-60°С), экстремальных термофилов (70 °С) и гипертермофилов (80 °С и выше). Споры отд. видов Б. выдерживают кратковрем. нагревание до 160-180 °C и длительное охлаждение до -196 °C и ниже. Некоторые Б. чрезвычайно устойчивы к ионизирующему излучению и живут даже в воде охлаждающих контуров атомных реакторов (Deinococcus radiodurans). Ряд Б. (барофилы, или пьезофилы) хорошо переносят гидростатич. давление до 101 тыс. кПа, а отд. виды не растут при давлении ниже 50 тыс. кПа. В то же время есть Б., не выдерживающие даже незначит. увеличения атмосферного давления. Большинство видов Б. не развиваются, если концентрация солей (NaCl) в среде превышает 0,5 моль/л. Оптимальные условия для развития умеренных и экстремальных галофилов наблюдаются в средах с концентрацией NaCl 10 и 30% соответственно; они могут расти даже в насыщенных растворах солей. Как правило, Б. предпочитают нейтральные условия среды обитания (рН ок. 7,0), хотя встречаются как экстремальные ацидифилы, способные к росту при рН 0,1-0,5, так и алкалифилы, развивающиеся при рН до 13,0. Подавляющее большинство изученных Б. - аэробы. Некоторые из них могут расти только при незначит. концентрации O 2 - до 1,0-5,0% (микроаэрофилы). Факультативные анаэробы растут как в присутствии O 2, так и в его отсутствие; они способны переключать метаболизм с аэробного дыхания на брожение или анаэробное дыхание (энтеробактерии). Рост аэротолерантных анаэробов не угнетается в присутствии небольшого количества O 2, т. к. они не используют его в процессе жизнедеятельности (напр., молочнокислые Б.). Для строгих анаэробов даже следы O 2 в среде обитания являются губительными. Многие Б. переживают неблагоприятные условия среды, образуя покоящиеся формы.Типы питанияДля Б. характерны интенсивный обмен веществами между клеткой и внешней средой и пластичность метаболизма. Они обладают высокой способностью к адаптации, легко приспосабливаясь к разл. (в т. ч. экстремальным) условиям среды, способны переключаться с одного типа питания на другой. Как и др. организмы, Б. запасают энергию гл. обр. в форме АТФ, образующегося в процессе фотосинтеза, дыхания и разл. типов брожения. В зависимости от источника используемого углерода они делятся на автотрофов (полностью удовлетворяют свои потребности за счёт CO 2) и гетеротрофов (нуждаются в готовых органич. соединениях). Однако эти термины не отражают всё многообразие типов питания у Б. Поэтому при их характеристике указывают на источник энергии, донор водорода (электронов) и вещества, используемые в биосинтетич. процессах. Для большинства Б. источником энергии служит окисление химич. веществ (хемотрофы). Ряд Б. (в т. ч. пурпурные и зелёные Б., цианобактерии) в ходе фотосинтеза преобразуют энергию света в энергию химич. связей органич. соединений (фототрофы). Если окислению подвергаются неорганич. вещества (т. е. они используются в качестве доноров водорода), Б. называют литотрофами (хемосинтетиками), если же вещества органич. природы, - органотрофами. Исходя из этого выделяют 8 типов питания и соответствующих им групп Б.: фотолитоавтотрофы (цианобактерии, анаэробные пурпурные Б.), фотолитогетеротрофы (некоторые анаэробные Б.), фотоорганогетеротрофы (несерные пурпурные Б.), фотоорганоавтотрофы (редкий тип питания, свойственный некоторым пурпурным Б.), хемолитоавтотрофы (напр., нитрификаторы, тионовые Б.), хемолитогетеротрофы (мн. сульфатвосстанавливающие Б.), хемоорганоавтотрофы (мн. водородные Б.), хемоорганогетеротрофы (осн. массив Б.-органотрофов). Известны облигатные паразиты (паратрофы), использующие только сложные органич. вещества, образуемые организмом-хозяином. Большинство Б., утилизирующих соединения азота, как правило, используют его восстановленные формы (чаще всего соли аммония), некоторые нуждаются в готовых аминокислотах, а другие усваивают и его окислённые формы (гл. обр. нитраты). Значит. число свободноживущих и симбиотических Б. способны фиксировать молекулярный азот (см. в ст. Азотфиксация). Фосфор, входящий в состав нуклеиновых кислот и др. соединений клетки, Б. получают преим. из фосфатов. Источником серы, необходимой для биосинтеза аминокислот и некоторых кофакторов ферментов, чаще всего являются сульфаты; некоторые виды Б. нуждаются в восстановленных соединениях серы.СистематикаОфициально принятой классификации Б. нет. Первоначально для этих целей использовалась искусств. классификация, основанная на сходстве их морфологич. и физиологич. признаков. Более совершенная филогенетическая (естественная) классификация объединяет родств. формы, исходя из общности их происхождения. Такой подход стал возможным после выбора в качестве универсального маркера гена 16S рРНК и появления методов определения и сравнения нуклеотидных последовательностей. Ген, кодирующий 16S рРНК (входит в состав малой субчастицы прокариотич. рибосомы), присутствует у всех прокариот, характеризуется высокой степенью консервативности нуклеотидной последовательности, функциональной стабильностью. Наиболее употребимой является классификация, публикуемая в периодич. издании определителя Бэрджи (Берги); см. также сайт в Интернете - http://141.150.157.117:8080/prokPUB/index.htm. По одной из существующих систем организмов, Б. вместе с археями составляют царство прокариот. Многие исследователи рассматривают их как домен (или надцарство), наряду с доменами (или надцарствами) архей и эукариот. В пределах домена наиболее крупными таксонами Б. являются филумы: Proteobacteria, включающий 5 классов и 28 порядков; Actinobacteria (5 классов и 14 порядков) и Firmicutes (3 класса и 9 порядков). Кроме того, выделяются таксономич. категории более низкого ранга: семейства, роды, виды и подвиды. По совр. представлениям, к одному виду относят штаммы Б., у которых последовательности нуклеотидов в генах, кодирующих 16S рРНК, совпадают более чем на 97%, а уровень гомологии нуклеотидных последовательностей в геноме превышает 70%. Описано не более 5000 видов Б., которые представляют лишь незначительную их часть среди населяющих нашу планету.Значение бактерийБ. являются самыми древними организмами, появившимися ок. 3,5 млрд. лет назад в архее. Ок. 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы. После появления многоклеточных организмов между ними и Б. образовались многочисл. связи, включая преобразование органич. веществ органотрофами, и разного рода симбиотич. отношения, паразитизм, иногда внутриклеточный (риккетсии), и патогенез. Наличие Б. и др. микроорганизмов в естеств. местах обитания является важнейшим фактором, определяющим целостность экологич. систем. В экстремальных условиях, непригодных для существования др. организмов, Б. могут представлять единственную форму жизни. Б. активно участвуют в биогеохимических циклах на нашей планете (в т. ч. в круговороте большинства химич. элементов). Совр. геохимич. деятельность Б. имеет также глобальный характер. Напр., из 4,3·10 10 т (гигатонн) органич. углерода, фиксированного в процессе фотосинтеза в Мировом океане, ок. 4,0 · 10 10 т минерализуется в водной толще, причём 70-75% из них - Б. и некоторыми др. микроорганизмами, а суммарная продукция восстановленной серы в осадках океана достигает 4,92 · 10 8 т в год, что почти в три раза превышает суммарную годовую добычу всех видов серосодержащего сырья, используемого человечеством. Осн. часть парникового газа - метана, поступающего в атмосферу, образуется Б. (метаногенами). Б. являются ключевым фактором почвообразования, зон окисления сульфидных и серных месторождений, образования железных и марганцевых осадочных пород и т. д. Некоторые Б. вызывают тяжёлые заболевания у человека, животных и растений. Нередко они становятся причиной порчи с.-х. продукции, разрушения подземных частей зданий, трубопроводов, металлич. конструкций шахт, подводных сооружений и т. д. Изучение особенностей жизнедеятельности этих Б. позволяет разработать эффективные способы защиты от вызываемых ими повреждений. В то же время положительную роль Б. для человека невозможно переоценить. С помощью Б. получают вино, молочные продукты, закваски и др. продукты, ацетон и бутанол, уксусную и лимонную кислоты, некоторые витамины, ряд ферментов, антибиотики и каротиноиды; Б. участвуют в трансформации стероидных гормонов и др. соединений. Их используют для получения белка (в т. ч. ферментов) и ряда аминокислот. Применение Б. для переработки с.-х. отходов в биогаз или этанол даёт возможность создания принципиально новых возобновляемых энергетич. ресурсов. Б. используют для извлечения металлов (в т. ч. золота), увеличения нефтеотдачи пластов (см. в статьях Бактериальное выщелачивание, Биогеотехнология). Благодаря Б. и плазмидам стало возможным развитие генетич. инженерии. Изучение Б. сыграло огромную роль в становлении мн. направлений биологии, в медицине, агрономии и др. Велико их значение в развитии генетики, т. к. они стали классич. объектом для изучения природы генов и механизмов их действия. С Б. связано установление путей метаболизма разл. соединений и др. Потенциал Б. в практич. отношении неисчерпаем. Углубление знаний об их жизнедеятельности открывает новые направления эффективного использования Б. в биотехнологии и др. отраслях промышленности.Литература Лит.: Шлегель Г. Общая микробиология. М., 1987; The Prokaryotes: Electronic release 3.0-3.17-. N. Y., 1999-2004-; Заварзин Г. А., Колотилова Н.Н. Введение в природоведческую микробиологию. М., 2001; Madigan M. T., Martinko J., Parker J. Brock biology of microorganisms. 10th ed. Upper Saddle River, 2003; Экология микроорганизмов. М., 2004. Барофильные микроорганизмыБАРОФИЛЬНЫЕ МИКРООРГАНИЗМЫАвторы: А.И. НетрусовБАРОФИЛЬНЫЕ МИКРООРГАНИЗМЫ, барофилы, пьезофилы (от греч. β ά ρος - тяжесть и φιλ έ ω - любить), организмы, способные расти при высоком давлении. Подразделяются на баротолерантные, умеренные и экстремальные. Баротолерантные Б. м. могут расти как при низких, так и при высоких значениях давления (до 40 МПа), умеренные барофилы растут в диапазоне 10-60 МПа, а экстремальные не развиваются при давлении ниже 40 МПа, максимальная скорость роста наблюдается у них при 70-81 МПа (иногда до 121 МПа). Б. м. обитают в глубоких слоях океана, часто встречаются в районах, связанных с активностью изотермальных источников (т. н. чёрных курильщиков); их выделяют также из глубоких нефтяных скважин. Представлены в осн. прокариотами - истинными бактериями и археями. |
 |