

Автор: Болдырев А.А.Статей: 2Биологические мембраны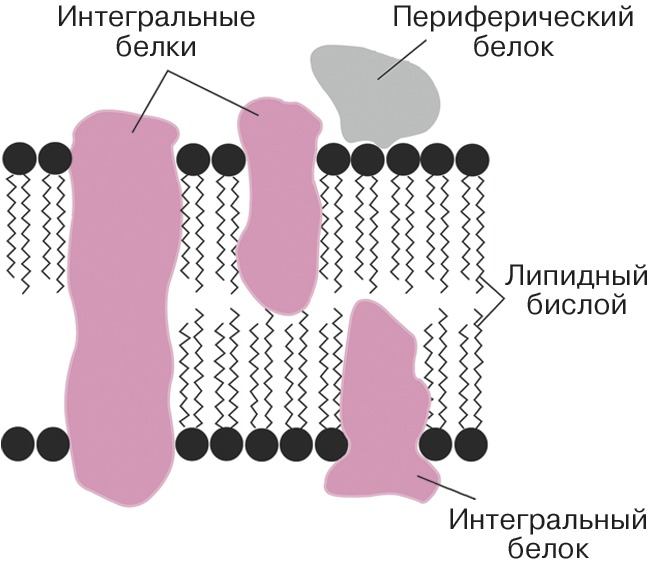 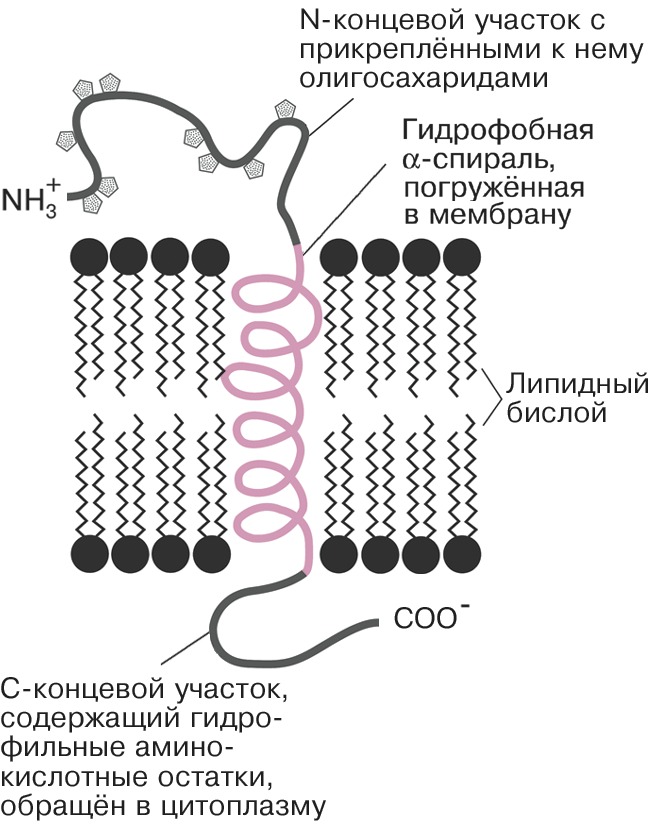 Схема строения клеточной мембраны. БИОЛОГИЧЕСКИЕ МЕМБРАНЫАвторы: А.А. БолдыревБИОЛОГИЧЕСКИЕ МЕМБРАНЫ (лат. membrana - кожица, оболочка, перепонка), структуры, ограничивающие содержимое клеток (клеточная, или плазматическая, мембрана, плазмалемма) и внутриклеточных органелл. У прокариот имеется только клеточная мембрана, в большинстве случаев окружённая клеточной стенкой. У эукариот мембраной окружена не только клетка, но и ядро, а также митохондрии, лизосомы, пероксисомы, секреторные гранулы, эндосомы, у растений ещё - хлоропласты и вакуоли; мембраны образуют также разветвлённую сеть эндоплазматического ретикулума и комплекса Гольджи. Митохондрии, хлоропласты и ядра окружены двумя мембранами, а внутри хлоропластов имеется ещё один тип мембран, формирующих тилакоиды. У животных к клеточной мембране снаружи примыкает гликопротеиновый комплекс - гликокаликс, у растений - клеточная стенка. Толщина мембран варьирует от 6 до 10 нм. Структура биологических мембранОснову Б. м. составляет протяжённый двойной слой (бислой) глицерофосфо-, сфинго- и гликолипидов со встроенными в него молекулами различных белков. Гидрофобные (неполярные) группы молекул липидов (остатки жирных кислот) погружены в толщу мембраны, а гидрофильные (полярные) головки ориентированы наружу, в окружающую водную среду (см. Липиды). Плотность упаковки Б. м. обеспечивается электростатическими взаимодействиями полярных головок и гидрофобными контактами между цепями жирных кислот. Входящие в состав Б. м. белки взаимодействуют с липидным бислоем с помощью гидрофобных взаимодействий и вандерваальсовых связей. Соотношение липидов и белков, их состав в разл. Б. м. могут существенно различаться. Так, в мембранах миелиновой оболочки содержание липидов (по массе) в четыре раза больше, чем белков, а во внутр. мембранах митохондрий более чем в два раза преобладают белки. Липиды Б. м. представлены гл. обр. фосфатидилхолином, фосфатидилэтаноламином, сфингомиелином, фосфатидилсерином, фосфатидилинозитом и кардиолипином, которые обнаруживаются примерно в одном и том же соотношении в мембранах разных по уровню организации организмов. В то же время набор жирных кислот, входящих в состав липидов, подвержен изменениям. Напр., понижение темп-ры, давления и солёности среды обитания организмов сопровождаются увеличением количества ненасыщенных связей и/или короткоцепочечных жирных кислот в фосфолипидах и происходящим вследствие этого уменьшением плотности упаковки бислоя. Липидам свойственна определённая подвижность внутри бислоя. Они способны к быстрому вращению вокруг оси (вращательная диффузия), к свободному перемещению в пределах одного слоя мембраны (латеральная диффузия), а также к переходу с одной стороны бислоя на другую (такое передвижение обеспечивается спец. механизмами). Для клеточных мембран животных клеток характерно высокое содержание холестерина (в ср. ок. 21%), который участвует в регуляции текучести мембраны, препятствуя плотной упаковке фосфолипидов. В растит. клетке роль холестерина играет его аналог - десмостерин. В мембранах бактерий и внутриклеточных органелл стерины отсутствуют. До 10% сухого вещества мембран приходится на долю углеводов, которые экспонированы на внешней стороне клеточной мембраны и являются составной частью мембранных гликолипидов и гликопротеинов. Содержание белка в разл. мембранах колеблется от 20 до 75% (в пересчёте на сухую массу). Мембранные белки могут быть встроены в бислой (интегральные белки). При этом они погружены в мембрану и пронизывают её (иногда неск. раз) таким образом, что достаточно протяжённые участки белка, образованные гидрофобными аминокислотами, оказываются в её толще, а гидрофильные - на поверхности, по обе Схема структурной организации интегрального белка. Спиральный участок встроен в гидрофобную часть липидного бислоя; N- концевой участок с прикреплёнными к нему олигосахаридными цепями расположен на вне... стороны Б. м. Выступающие над внешней стороной мембраны участки белковых молекул обычно несут неск. ковалентно связанных, часто разветвлённых цепей олигосахаридов, образованных остатками маннозы, фукозы, глюкозы, N-ацетилглюкозамина и др. Эти компоненты играют роль маркеров при распознавании клеточной поверхности. Молекулы периферических белко в расположены гл. обр. на внутренней поверхности мембраны, не проникая внутрь бислоя, и удерживаются на ней с помощью электростатич. взаимодействий и водородных связей; они связываются с мембраной обратимо и могут переходить в цитоплазму при модификации белков (напр., путём их фосфорилирования) в ответ на изменения функционального состояния клетки. Мн. белки организованы в виде сложных комплексов (напр., белки дыхательной цепи митохондрий). В клетках происходит постоянное обновление компонентов Б. м. путём введения новых молекул липидов и белков, однако структурная организация Б. м. в течение всей жизни клетки остаётся неизменной.Функции мембранОсн. функции мембран связаны с белками. Мн. мембранные белки - ферменты, обеспечивающие протекание окислительно-восстановительных, гидролитич. и биосинтетич. реакций как на поверхности мембраны, так и внутри неё. Важнейшая функция белков мембран - транспортная. Жирорастворимые соединения (напр., стероидные гормоны) легко проникают в липидный бислой, но для большинства др. соединений (в т. ч. аминокислот, сахаров) и неорганич. ионов он непроницаем. Специально предназначенные для этих целей мембранные белки обеспечивают как активный (требующий затрат энергии), так и пассивный (за счёт градиента концентраций) транспорт веществ и ионов (см. Ионные каналы). Специфич. белки - ионные насосы (Na / K -насос и Ca -насос) ответственны за асимметричное распределение ионов Na +, K + и Ca 2 + по обе стороны клеточной мембраны (напр., в цитоплазме животной клетки - низкую для Na + и Ca 2 + и высокую для K +). Такая асимметрия обеспечивает мн. проявления жизнедеятельности (электровозбудимость, осмотическую устойчивость и др.). Мембранные белки аквапорины образуют в мембране спец. каналы, регулирующие проникновение в клетку молекул воды. Взаимодействие клетки с внешней средой, регуляция внутриклеточных процессов осуществляются посредством рецепторных белков (рецепторов), ответственных за фото-, термо-, механо- и хеморецепцию. Барьерная функция Б. м. обеспечивает сохранение определённого состава клетки и концентрации составляющих её веществ, а также защиту от воздействия разл. чужеродных факторов и токсинов. Благодаря Б. м. внутри клеток возможно одноврем. протекание множества несовместимых друг с другом реакций. Напр., необходимые клетке белки синтезируются на рибосомах, прикреплённых к эндоплазматическому ретикулуму, а их распад происходит в лизосомах. В Б. м. протекают процессы энергообмена клеток. Внутр. мембраны митохондрий и мембраны тилакоидов - важнейшие преобразователи энергии, играющие ключевую роль в запасании энергии, образующейся в ходе дыхания и фотосинтеза, в энергию пирофосфатной связи аденозинтрифосфата. Б. м. нейронов могут генерировать и осуществлять передачу электрич. сигнала, участвуя тем самым в процессах возбуждения и проведения нервного импульса. Белковые и липидные компоненты выполняют ряд др. функций. Фрагменты фосфолипидов могут выступать в качестве предшественников сигнальных молекул (мессенджеров). Напр., при активации мембранной фосфолипазы А из бислоя высвобождается арахидоновая кислота, дальнейшие превращения которой приводят к образованию биологич. регуляторов - тромбоксанов, лейкотриенов и простагландинов. Фосфатидилсерин, локализованный на внутр. стороне мембраны, при инициации апоптоза мигрирует на её внешнюю сторону. Его появление служит сигналом для фагоцитов, которые имеют рецепторы на этот фосфолипид; они «узнают» дефектные клетки и уничтожают их. Гликолипиды наряду с гликопротеинами играют важную роль в явлениях межклеточной адгезии, участвуют в иммунных реакциях.Методы изучения биологических мембранДля изучения структуры и функции мембран используются электрофизиологич. и иммуноцитохимич. методы, жидкостная хроматография (для идентификации и анализа липидных компонентов), проточная цитометрия, позволяющая проследить ответ клетки на взаимодействие специфич. лигандов с клеточной мембраной, разнообразные физич. методы, характеризующие структуру мембран, упаковку и подвижность липидов в бислое (в т. ч. электронная микроскопия, малоугловое рассеивание нейтронов, флуоресцентная спектроскопия, круговой дихроизм), и др. методы. Разнообразие типов Б. м., их полифункциональность и высокая чувствительность к внешним воздействиям являются причиной того, что они вовлекаются в разл. патологич. процессы. Повреждения клеточных мембран, приводящие к образованию свободных радикалов и гибели нервных клеток, лежат в основе нейродегенеративных заболеваний (болезнь Альцгеймера, паркинсонизм, боковой амиотрофический склероз), могут служить причиной инсульта и инфаркта миокарда.Литература Лит.: Крепс Е.М. Липиды клеточных мембран. Л., 1981; Введение в биомембранологию. М., 1990; Биологические мембраны. Методы. М., 1990; Геннис Р. Биологические мембраны. М., 1999; Пальцев М. А., Иванов А. А., Северин С.Е. Межклеточные взаимодействия. 2-е изд. М., 2003; Ченцов Ю.С. Введение в клеточную биологию. 4-е изд. М., 2004. КарнозинКАРНОЗИНАвторы: А.А. БолдыревКАРНОЗИН, дипептид, образованный остатками β -аланина и гистидина; присутствует в значит. количествах (0,4-2 г на 1 кг сырой массы) в составе головного и спинного мозга, сердечной и скелетной мышц позвоночных животных. Впервые обнаружен и описан В.С. Гулевичем в 1900, первые доказательства его биологич. активности (увеличение сократительной способности утомлённой мышцы) получены в 1953 С.Е. Севериным. Функция К. в организме обусловлена гл. обр. буферными свойствами, позволяющими препятствовать изменению pH, концентрации катионов металлов переменной валентности, а также свободных радикалов в клетке. В организме содержание К. регулируется ферментами карнозинсинтазой и карнозиназой, участвующими соответственно в его синтезе и распаде; он может превращаться в др. биологически активные соединения - анзерин, карцинин, гомокарнозин, ацетилированные производные. К. используется для лечения катаракты, в качестве ранозаживляющего и иммуномодулирующего препарата. Показана его способность восстанавливать работоспособность после физич. и эмоциональных перегрузок, на стадии реабилитации после разл. заболеваний, при радиотерапии злокачественных опухолей. Литература Лит.: Болдырев А.А. Карнозин. М., 1998; он же. Карнозин и защита тканей от окислительного стресса. М., 1999; Карнозин и окислительный стресс. М.; Петрозаводск, 2006. |
 |