

Автор: Ченцов Ю.С.Статей: 7ВакуолиВАКУОЛИАвторы: Ю.С. ЧенцовВАКУОЛИ, полости в цитоплазме клеток эукариот, а также некоторых прокариот, ограниченные одинарной липопротеиновой мембраной. Могут быть заполнены как жидкими (растворы белков и сахаров), так и плотными (кристаллы солей, нерастворимые белки) компонентами. В клетках эукариот В. имеют вид сферич. пузырьков (везикулы), плоских мешков (цистерны) или узких ветвящихся канальцев и объединены в т. н. вакуолярную систему. В клетках прокариот В. являются производными плазматич. мембраны, от которой в цитоплазму клетки отщепляются мембранные пузырьки. Они характерны для некоторых фотосинтезирующих бактерий (в т. ч. цианобактерий), в В. (хроматофорах, тилакоидах) которых локализованы компоненты фотосинтетич. системы. У мн. видов водных фототрофных и хемотрофных бактерий присутствуют газовые В. (аэросомы). Последние выполняют роль регуляторов плавучести, позволяя обладающей ими клетке занимать в слое воды оптимальное положение относительно интенсивности света, концентрации растворённого кислорода или содержания питательных веществ. Вакуолярная системаВАКУОЛЯРНАЯ СИСТЕМААвторы: Ю.С. ЧенцовВАКУОЛЯРНАЯ СИСТЕМА, форма организации вакуолей в клетках эукариот. В. с. состоит из эндоплазматической сети (эндоплазматич. ретикулума), ядерной оболочки, комплекса Гольджи, лизосом и многих др. вакуолей. В ней происходят синтез, модификация, сортировка и выделение из клетки биополимеров, гл. обр. белков (ферменты, гликопротеины), синтез биологич. мембран, в т. ч. плазматической. Последняя как производная этой системы может образовывать за счёт инвагинации (впячивания внутрь клетки) вторичные вакуоли - эндосомы, участвующие в поглощении внеклеточных компонентов (пиноцитозные и фагоцитозные вакуоли). Эндосомы, объединяясь с первичными лизосомами, содержащими гидролитич. ферменты, превращаются в вакуоли, выполняющие функцию внутриклеточного пищеварения (вторичные лизосомы). Для В. с. характерны кооперативность функционирования, взаимосвязь и последовательность образования, транспорта и выделения синтезиров. белков. Все компоненты В. с. имеют единый источник образования - гранулярную (шероховатую) эндоплазматич. сеть. На её рибосомах происходит биосинтез мембранных и внутривакуолярных секреторных белков. Затем от мембран сети отделяются мелкие вакуоли, содержащие растворимые белки, и переносятся в зону мембран комплекса Гольджи, где они претерпевают ряд сложных изменений. В дальнейшем вакуоли, наполненные секреторными белками, отделяются от мембран комплекса Гольджи, подходят к плазматич. мембране, сливаются с ней, а их содержимое выводится из клетки (секретируется). Кроме того, от гранулярной эндоплазматич. сети отщепляются трубчатые мембранные компоненты - гладкие эндоплазматич. сети, которые участвуют в биосинтезе липидов и полисахаридов. В клетках растений, кроме описанных выше вакуолей В. с. (исключение составляют фагоцитарные вакуоли, свойственные клеткам животных), важную роль играют т. н. центральные вакуоли. Они отделены от цитоплазмы липопротеиновой мембраной (тонопласт), сходной с плазматической. Заполняющая их жидкость представляет собой концентрир. раствор, содержащий растворённые в воде соли, сахара, аминокислоты, белки, конечные продукты обмена веществ (таннины, гликозиды, алкалоиды) и некоторые пигменты (напр., антоцианы). Эти вакуоли участвуют в регуляции водно-солевого обмена, поддержании тургорного давления в растит. клетках, накоплении низкомолекулярных водорастворимых веществ и запасных веществ, а также в выведении токсичных продуктов. Литература Лит.: Фаллер Д. М., Шилдс Д. Молекулярная биология клетки. М., 2003; Ченцов Ю.С. Введение в клеточную биологию. М., 2004; Molecular cell biology. 5th ed. N. Y., 2004. Интерстициальные клеткиИНТЕРСТИЦИАЛЬНЫЕ КЛЕТКИАвторы: Ю.С. ЧенцовИНТЕРСТИЦИАЛЬНЫЕ КЛЕТКИ (от лат. interstitium - промежуток), общее название разл. клеток животных и человека, расположенных, как правило, между осн. элементами, формирующими тот или иной орган и выполняющими главную функциональную нагрузку. Некоторые И. к. являются продуцентами гормонов (напр., клетки, находящиеся между семенными канальцами в семенниках, вырабатывают мужские половые гормоны, гл. обр. тестостерон; И. к. яичников продуцируют ряд стероидных гормонов). И. к. мозгового вещества почек участвуют в поддержании солевого и водного гомеостаза. У книдарий И. к. (i-клетки) могут развиваться в половые, нервные или стрекательные клетки и участвовать в процессах регенерации. Гольджи аппарат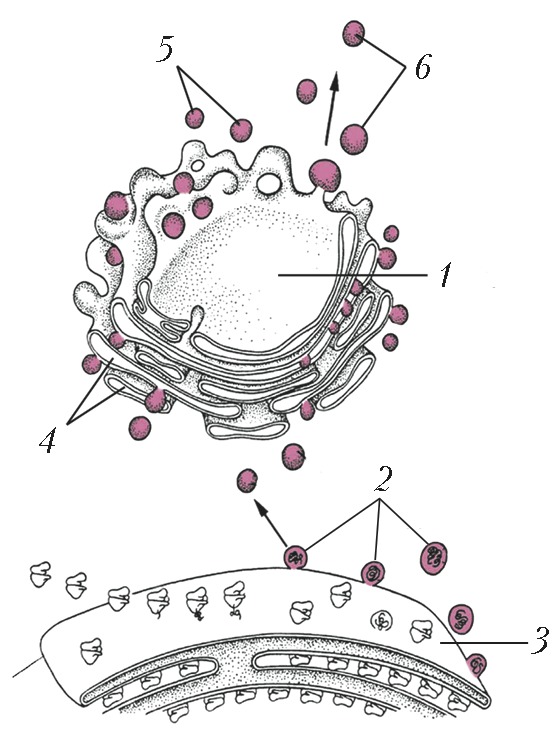 Гольджи аппарат: 1 - диктиосома; 2 - вакуоли, отделяющиесяот эндоплазматической сети;3 - эндоплазматическая сеть; 4 - цистерны; 5 - секреторные вакуоли; 6 - вакуоли... ГОЛЬДЖИ АППАРАТАвторы: Ю.С. ЧенцовГОЛЬДЖИ АППАРАТ (комплекс Гольджи), клеточная органелла, часть вакуолярной системы клетки. Названа в честь К. Гольджи, который в 1898, рассматривая под микроскопом кусочки ткани головного мозга, предварительно обработанные солями тяжёлых металлов, обнаружил структуру, названную им «сетчатым аппаратом клетки». Впервые предложения об участии этой структуры в выведении из клетки разл. продуктов высказаны Д.Н. Насоновым (1923). Позднее с помощью электронного микроскопа было показано присутствие Г. а. во всех эукариотич. клетках (кроме эритроцитов и зрелых сперматозоидов), установлены его строение и механизм действия. Функциональной единицей Г. а. является диктиосома (рис.), состоящая из стопки уплощённых, окружённых липопротеиновой мембраной вакуолей, или цистерн (до 20). Диктиосома всегда окружена множеством ограниченных мембраной сферич. вакуолей. Весь Г. а. клетки обычно включает неск. диктиосом (до 10), связанных друг с другом каналами и образующих неупорядоченную сетчатую структуру, чаще около ядра. В клетках некоторых типов (в т. ч. клетках нервного ганглия) компоненты Г. а. бывают рассеяны по всей цитоплазме. Образование и работа Г. а. целиком зависят от гранулярной (шероховатой) эндоплазматической сети (ЭС). К ЭС обращена одна из поверхностей диктиосомы, называемая формирующейся проксимальной, или цис -зоной (от лат. cis - по эту сторону, вблизи). Другая поверхность, которая называется зрелой дистальной, или транс -зоной (от лат. trans - по ту сторону), ориентирована в сторону цитоплазматич. мембраны. Кроме того, выделяют расположенную между ними среднюю зону. В гранулярной ЭС синтезируются как растворимые (секреторные), так и нерастворимые, встраивающиеся в её мембрану белки (гликопротеины); в составе мелких вакуолей они отделяются от ЭС и сливаются с мембранами цис -зоны. В свою очередь, от этой зоны Г. а. отщепляются вакуоли, переносящие секреторные и мембранные белки в среднюю зону, а от последней - в транс -зону. В Г. а. происходит вторичная модификация белков, поступающих после их синтеза на рибосомах гранулированной ЭС: в цис -зоне - фосфорилирование гидролаз и некоторых мембранных белков; в средней - вторичное гликозилирование белков, их окончат. «созревание»; в транс -зоне, помимо модификации белков, происходит сортировка растворимых белков, их накопление в отд. вакуолях за счёт взаимодействия специфически модифициров. участков с рецепторными белками в составе транс -зон Г. а. При этом растворимые белки разделяются на три осн. группы: белки лизосом (гл. обр. гидролазы), белки постоянной секреции и белки регулируемой (сигнальной) секреции. Вакуоли Г. а., содержащие гидролазы, становятся лизосомами и участвуют во внутриклеточных процессах расщепления поглощённых в ходе эндоцитоза биополимеров, а также в деградации дефектных цитоплазматич. компонентов. Вакуоли, содержащие секреторные белки, транспортируются к плазматич. мембране (при участии микротрубочек цитоскелета) и сливаются с ней, выбрасывая своё содержимое из клетки (экзоцитоз) и одновременно пополняя её липопротеиновую мембрану. Литература Лит.: Уэйли У. Аппарат Гольджи. М., 1978; Molecular biology of the cell. 4th ed. N. Y., 2002; Molecular cell biology. 5th ed. N. Y., 2003; Ченцов Ю.С. Введение в клеточную биологию. 4-е изд. М., 2005. Клетка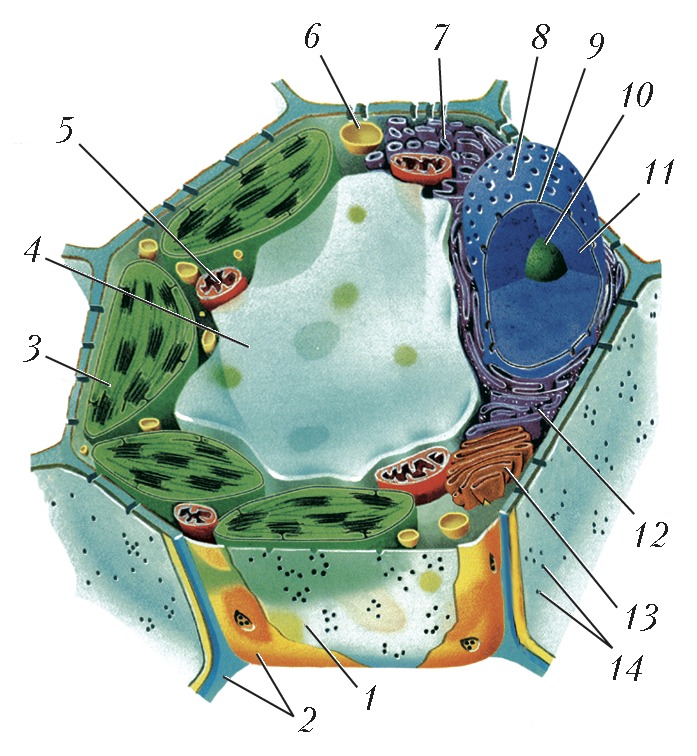 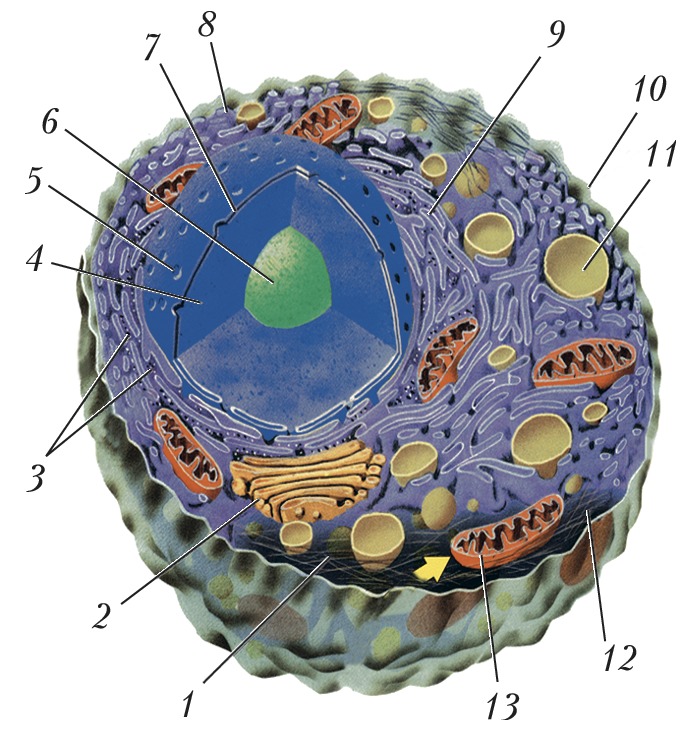 Схема строения растительной клетки: 1 - плазматическая мембрана; 2 - клеточная стенка; 3 - хлоропласты; 4 - вакуоль; 5 - митохондрия; 6 - лизосома;
КЛЕТКААвторы: Ю.С. ЧенцовКЛЕТКА (лат. cellula, греч. ϰύτος), основная структурно-функциональная единица всех живых организмов. Представляет собой сложноорганизованную систему, состоящую из биополимеров (в т. ч. нуклеиновых кислот, белков) и др. соединений, способную к самоподдержанию и самовоспроизведению. К. может существовать как самостоятельный одноклеточный организм (бактерии, археи, простейшие, некоторые водоросли и грибы), так и в составе тканей и органов разл. многоклеточных организмов. Историческая справкаТермин «К.» был предложен Р. Гуком (1665) для обозначения ячеек, которые он наблюдал с помощью увеличительных линз в срезах мёртвой пробковой ткани, представлявших собой лишь внешние оболочки растительных К. Позднее аналогичные «К.» Гука были обнаружены в составе разных частей растений и в тканях животных. Только в нач. 19 в. в связи с развитием микроскопии изменились общие представления о К.: главным в их организации стали считать не внешние оболочки («ячейки»), а содержимое - протоплазму (Я. Пуркине). В 1831 Р. Броун обнаружил и описал в протоплазме растит. К. постоянную структуру - ядро. На основании этих и многих др. наблюдений Т. Шванн сформулировал осн. положения клеточной теории (1839), одно из которых постулировало сходство строения К. растений и животных. Дальнейшее развитие клеточной теории связано с работами Р. Вирхова, доказавшего, что количество К. в организме увеличивается в результате клеточного деления (это правило было сформулировано им в его афористич. формуле - «Всякая клетка от клетки»). Создание клеточной теории явилось важнейшим событием в естествознании, одним из решающих доказательств единства всей живой природы. Основные её положения сохранили своё значение и в наше время. Совр. клеточная теория рассматривает К. как элементарную единицу живого; вне её нет жизни; увеличение числа К. происходит только путём деления исходных К.; многоклеточные организмы представляют собой сложные ансамбли К., объединённых в системе тканей и органов, и связаны межклеточными взаимодействиями; К. способны к метаболизму, использованию и трансформации энергии, чувствительности, изменчивости. Совокупность всех этих признаков можно обнаружить только на клеточном уровне.Типы организации клетокСреди живых организмов встречаются два типа строения К. — прокариотный и эукариотный. Прокариотные К. характерны для всех бактерий и архей; их размеры невелики (0,1-10 мкм); обычно это единичные, свободноживущие организмы, но некоторые из них могут образовывать колонии из одинаковых К. Они не имеют морфологически выраженного ядра - их геном (нуклеоид) в виде компактно упакованной кольцевой молекулы ДНК (т. н. бактериальной хромосомы) не отделён от цитоплазмы и структурно связан с плазматической мембраной, отграничивающей К.; снаружи прокариотная К. окружена клеточной стенкой (у архей т. н. S-слоями либо оболочкой, сходной с клеточной стенкой грамположительных бактерий); у некоторых прокариот клеточная стенка отсутствует. В цитоплазме (протоплазме) К. имеются вакуоли, многочисл. ферменты, обеспечивающие метаболизм, рибосомы, элементы цитоскелета; в К. могут находиться также и внехромосомные генетич. элементы - ДНК плазмид. Деление прокариотной К. — бинарное (путём образования перегородки), происходит после репликации ДНК. Эукариотные К. свойственны всем остальным организмам. Они в 10-100 раз крупнее прокариотных; содержат морфологически выраженное ядро, отграниченное от цитоплазмы мембранной ядерной оболочкой и представляющее собой сложную систему, обеспечивающую хранение, воспроизведение и реализацию генетич. информации, содержащейся в линейных молекулах ДНК, заключённых в хромосомах. В цитоплазме, ограниченной плазматич. мембраной, выделяют жидкую плазму (гиалоплазма, или цитозоль), участвующую в промежуточном метаболизме К., а также множество спец. обязательных (наряду с ядром) структурных образований (органелл), каждая из которых выполняет определённые специфич. функции (сходные у разл. эукариотных К.). Рибосомы осуществляют синтез белка, митохондрии обеспечивают К. энергией, пластиды растительных К. (в т. ч. хлоропласты) участвуют в синтезе АТФ и фотосинтезе. Митохондрии и хлоропласты содержат собственный генетич. аппарат, что послужило основанием для возникновения гипотезы симбионтного происхождения эукариотных К. (см. Симбиогенез). Часть органелл, окружённых мембраной [эндоплазматическая сеть (эндоплазматич. ретикулум), аппарат Гольджи, эндо- и экзоцитозные вакуоли, лизосомы, пероксисомы], образуют вакуолярную систему. Кроме того, для К. эукариот характерно наличие сети белковых нитей, пронизывающих цитоплазму, — цитоскелет (опорно- двигательная система К.). У К. растений и грибов снаружи плазматич. мембраны расположена клеточная стенка, отсутствующая в К. животных. Обязательным компонентом цитоплазмы К. животных является клеточный центр, в состав которого входит центриоль. Соматические К. эукариот делятся путём митоза, сопровождающегося образованием спец. аппарата деления - клеточного веретена, с помощью которого происходит равномерное распределение хромосом (после их удвоения) строго по двум дочерним клеткам, в которых сохраняется исходный диплоидный набор хромосом. Продолжительность жизни К. от деления до деления составляет клеточный цикл. Половые К. (гаметы) у растений и животных формируются в ходе мейоза (в каждой К. оказывается гаплоидный набор хромосом).Сходство и разнообразие клетокНесмотря на мн. морфологич. и функциональные особенности, прокариотные и эукариотные К. во многом сходны, что обусловлено общностью структурно-молекулярной организации их внутриклеточных структур, которые образуют в К. некое сопряжённое единство и функционально взаимозависимы: и прокариотные, и эукариотные К. окружены плазматич. мембраной, выполняющей барьерную, транспортную и рецепторную функции; оба типа К. участвуют в процессах, связанных с поддержанием самой живой системы (напр., синтеза нуклеиновых кислот и белков, в биоэнергетических). Размножение К. происходит только путём деления исходной клетки, причём прокариотные К. используют аппарат деления К., напоминающий митотический. Одновременно (при сходстве в строении внутриклеточных структур) эукариотные К. многоклеточных организмов могут различаться по форме, строению и размеру (даже в пределах одной особи, напр. К. эпителия, мышц, нервной системы, крови). Такое разнообразие К. можно объяснить специализацией функций, выполняемых разл. К., что является результатом избирательной активности разных генов в разных К. по мере развития многоклеточного организма (см. Дифференцировка). Показано, что любая К. такого организма обладает одинаковым полным набором генетич. материала, всеми возможностями для работы любого гена (т. е. полипотентна), но в разных К. одни и те же гены могут находиться в активном или в репрессивном состоянии. Всегда активны гены, определяющие синтез белков и нуклеиновых кислот, необходимых для поддержания жизни самой К., но в К. разных тканей, кроме того, активны специфич. гены, определяющие свойства и осн. клеточные функции, характерные именно для К. данной ткани. Регуляция же активности спец. генов определяется межклеточными (см. Межклеточные взаимодействия), гормональными, нервными (для животных) формами химич. регуляции. Т. о., в многоклеточном организме К. является единицей развития, единицей функционирования в разл. органах и тканях и первичным объектом патологич. изменений жизнедеятельности. У эукариот развитие организмов начинается с родоначальной К. (зиготы), потомки которой образуют сложные многоклеточные ансамбли тканей и органов. Любые функции многоклеточного организма - результат работы отд. К. в разных органах. Нарушение целостности К. или изменение их свойств - осн. причина всех без исключения заболеваний. Естественное разрушение клеток происходит в ходе реализации генетически детерминированной программы - апоптоза. Наука о К. — клеточная биология. Для изучения К. используют разл. виды микроскопии, культивирование К. (см. Культура клеток и тканей), разл. методы клеточной инженерии, а также многочисл. методы биохимии, биофизики, молекулярной биологии.Литература Лит.: Вирхов Р. Патология, основанная на теории ячеек, (целлулярная патология) в применении к микроскопической анатомии нормальных и ненормальных тканей. М., 1859; Кацнельсон З.С. Сто лет учения о клетке. М.; Л., 1939; Вермель Е.М. История учения о клетке. М., 1970; Александров В.Я. Реактивность клеток и белки. Л., 1985; Ченцов Ю.С. Введение в клеточную биологию. М., 2004; Pollard T. D., Earnshaw W. C. Cell biology. 2nd ed. Phil., 2008. См. также лит. при ст. Клеточная биология. Клеточная биологияКЛЕТОЧНАЯ БИОЛОГИЯАвторы: Ю.С. ЧенцовКЛЕТОЧНАЯ БИОЛОГИЯ, наука о физико-химич. основах строения клетки, структуре, функции её мембранных компонентов, о работе её генетич. аппарата, механизмах и путях внутриклеточного перемещения белков и роли в этих процессах мембран и цитоскелета, о принципах структурной организации обменных процессов, о сигнальных механизмах внутри- и межклеточных взаимодействий; предметом изучения К. б. являются также особенности жизненного цикла клеток и его регуляции, молекулярные и структурные основы митоза и мейоза, а также особенности репарации, структурные и молекулярно-биологич. изменения клеток при их необратимых повреждениях и запрограммированной клеточной смерти (апоптозе). Впервые термин «К. б.» использован в 1884 франц. учёным Ж.Б. Карнуа в монографии, обобщающей сведения о клетке («La biologie cellulaire»). Однако вплоть до сер. 20 в. в отеч. науч. лит-ре для определения морфологич. науки о строении клетки употреблялся термин цитология, осн. методом исследования которой была световая микроскопия, позволившая получить важнейшие сведения о компонентах клеток и их функциях. С 1950-х гг. благодаря успехам физики и химии в цитологии начали применяться методы, давшие возможность изучать клетку также на молекулярном уровне. Именно в этот период возродился термин «К. б.», который отражал формирование новой комплексной синтетич. науки о клетке. Объединение усилий и интересов исследователей классич. биохимии, молекулярной биологии, генетики и собственно цитологии, использующих электронную микроскопию для изучения субклеточных структур, новые методы молекулярной биологии, иммунохимии, молекулярно-генетич. анализа и др., позволило раскрыть пути межклеточных взаимоотношений в многоклеточных организмах и особенности специализации (дифференцировки) разл. клеток. В связи с применением приёмов молекулярной биологии для решения широкого круга вопросов К. б. её часто называют молекулярной клеточной биологией. Современная К. б., объединяющая мн. науч. направления, не только приблизилась к завершению изучения физиологии клетки как таковой, но и сохранила и укрепила осн. положения классич. цитологии и клеточной теории. Достижения К. б. широко используются для решения общебиологич. задач, в медицине, сельском хозяйстве. Литература Лит.: Молекулярная биология клетки. 2-е изд. М., 1994. Т. 1-3; Molecular biology of the cell. 4th ed. N. Y., 2002; Molecular cell biology. 5th ed. N. Y., 2003. ЦитологияЦИТОЛОГИЯАвторы: Ю.С. ЧенцовЦИТОЛОГИЯ (от цито... и...логия), наука о клетке. Изучает строение и функции тканевых клеток у многоклеточных организмов, одноклеточные организмы и ядерно- цитоплазматич. комплексы, не расчленённые на клетки (симпласты, синцитии, плазмодии). Клетка является элементарной структурой, определяющей строение, функционирование и развитие организмов. Поэтому данные Ц. широко используются в самых разных областях биологии, в т. ч. в гистологии, анатомии, эмбриологии, физиологии, генетике, молекулярной биологии. Изучение клеток началось во 2-й пол. 17 в. благодаря изобретению и использованию микроскопа. Клеточное строение в ряде растит. тканей впервые наблюдал Р. Гук (1665). Позднее появились работы Н. Грю, М. Мальпиги, А. ван Левенгука, которые показали, что разнообразные части растений состоят из ячеек, или клеток. Левенгук, кроме того, впервые увидел клетки животных и микроорганизмы. Прогресс изучения клетки связан с развитием техники микроскопирования. В 19 в. описание Р. Броуном клеточного ядра (1831) привлекло внимание исследователей к содержимому клетки. К 1839 была создана клеточная теория (Т. Шванн) - одно из важнейших обобщений в биологии. Во 2-й пол. 19 в. открыты постоянные составные части цитоплазмы (органеллы) и выявлена роль ядра в процессе деления клеток. В нач. 20 в. изучение роли хромосом в наследственности и определении пола привело к формированию цитогенетики. Была разработана методика культивирования тканевых клеток вне организма, что является неотъемлемой частью технологии культивирования тканей и тканевой инженерии, начаты цитохимич. исследования клеток. Новый этап развития Ц. (с 1950-х гг.) связан с использованием электронного микроскопа, методов количественной цитохимии, включая авторадиографию, и др. химич. и физич. методы исследования при непосредств. взаимодействии с биохимией и молекулярной биологией. Это привело к выявлению ряда новых структур в цитоплазме и ядре и позволило расшифровать их функциональное значение. Именно в этот период появилась новая комплексная наука - клеточная биология. В Ц. существует ряд направлений, решающих частные задачи: кариосистематика, радиационная Ц., иммуноцитология, цитопатология и др. Важное достижение в развитии Ц. - создание совокупности методов, позволяющих конструировать клетки, - клеточной инженерии. См. также Клетка. Литература Лит.: Вермель Е.М. История учения о клетке. М., 1970; Molecular biology of the cell. 4th ed. N. Y.; L., 2002; Molecular cell biology. 5th ed. N. Y., 2003; Ченцов Ю.С. Введение в клеточную биологию. 4-е изд. М., 2005; Pollard T. D., Earnshaw W. C. Cell biology. 3rd ed. Phil., [2017]. |
 |