

И > ИМ Иммуноглобулин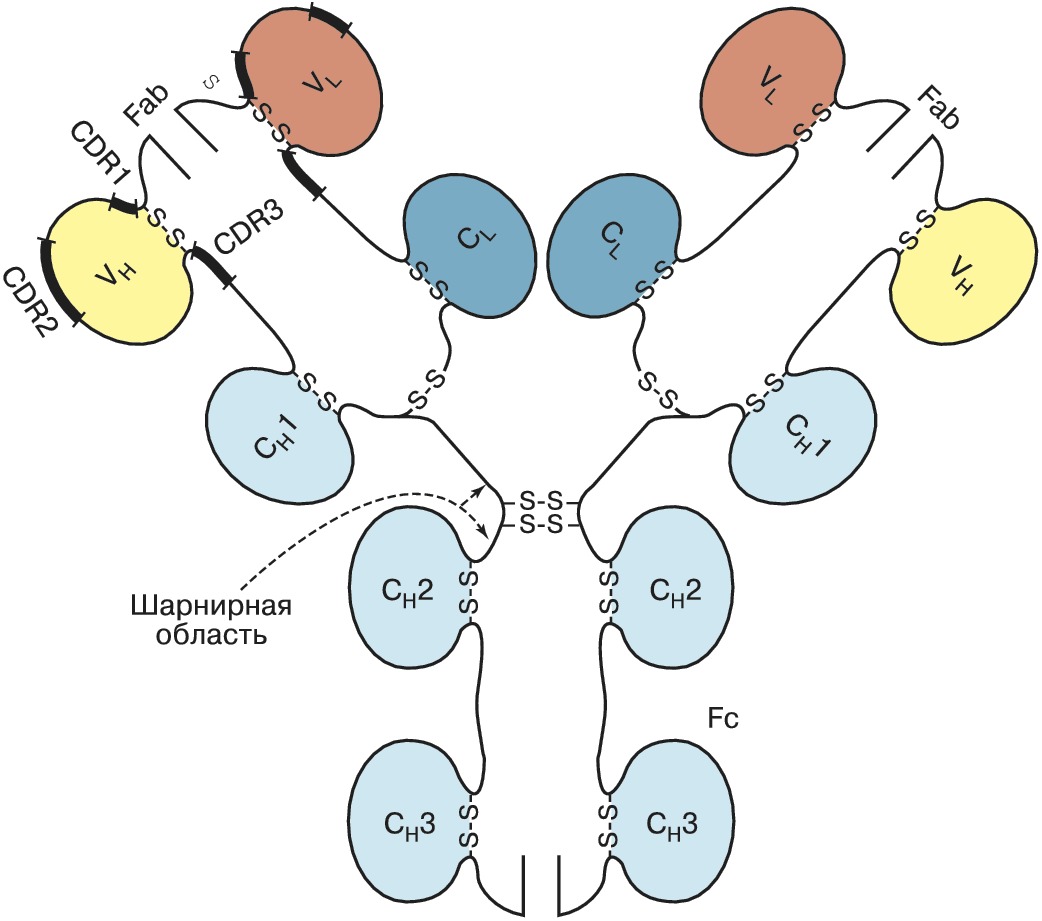 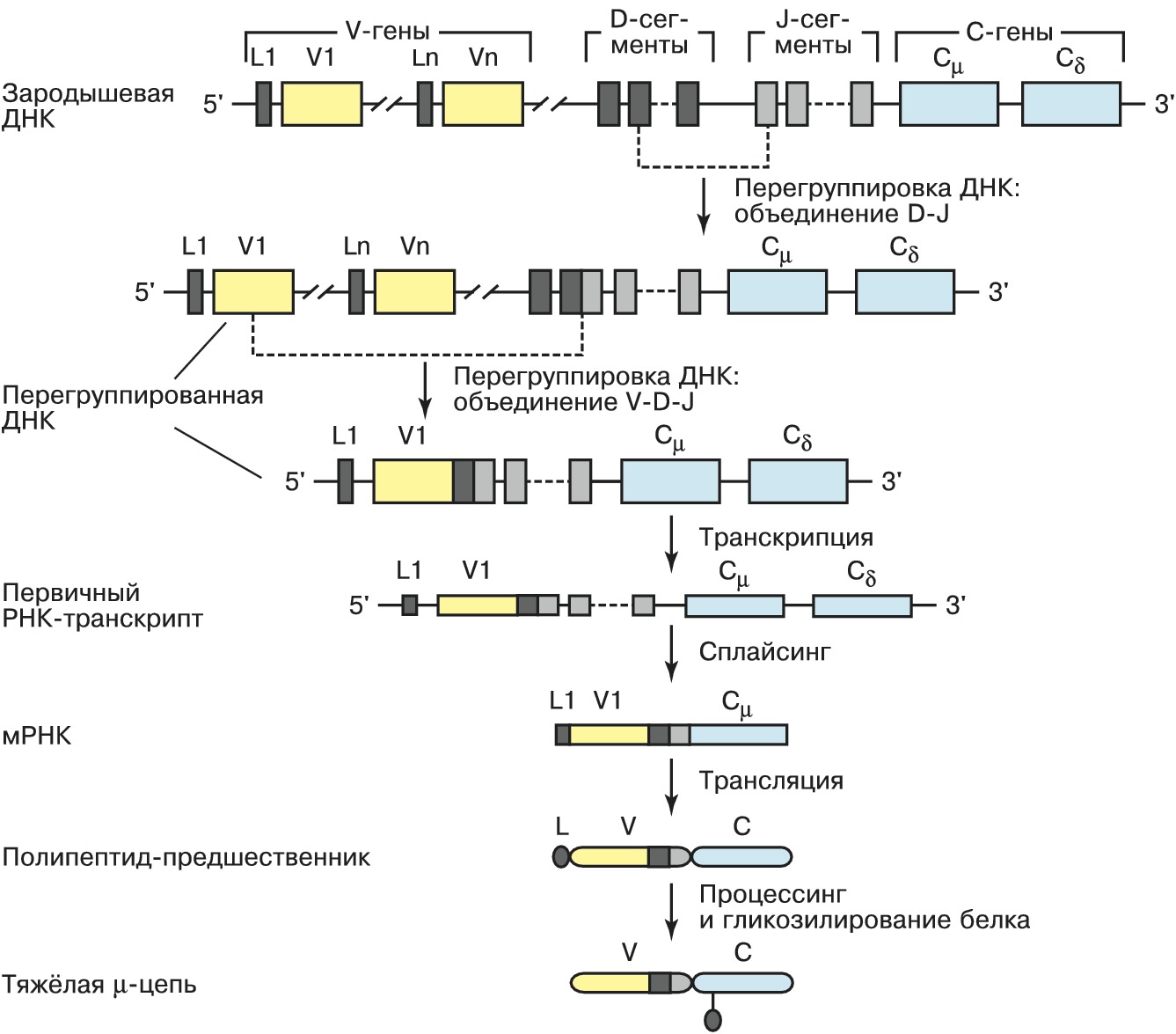 Рис. 1. Строение молекулы IgG1 человека: VH и VL - вариабельные домены соответственно тяжёлой и лёгкой цепей; CL - константный домен лёгкой цепи; СH1, СH2, СH3 - константные домены ИММУНОГЛОБУЛИНАвторы: С.М. ДеевИММУНОГЛОБУЛИН (Ig), сложный глобулярный белок позвоночных животных (начиная с хрящевых рыб) и человека, обладающий активностью антитела, т. е. способностью специфически взаимодействовать с антигеном (см. Антиген - антитело реакция). И. присутствует либо в свободном виде в крови и др. жидкостях организма, либо в виде рецепторов на поверхности клеточных мембран. У каждого индивидуума существует громадное разнообразие молекул И., различающихся по специфичности. Они синтезируются плазматич. клетками, дифференцировка которых инициируется взаимодействием В-лимфоцитов иммунной системы (имеющих рецептор нужной специфичности) с антигенами при участии ряда медиаторов (напр., интерлейкинов). Уникальная специфичность И. и высокое сродство к «своему» антигену обеспечивается комплементарностью активного центра И. к участку молекулы антигена (эпитопу). Одновременно И. могут связываться с рецепторами лейкоцитов (т. н. Fc-рецепторами) или белками системы комплемента, в результате чего включается цепь последовательных реакций иммунной системы организма, приводящих к удалению чужеродного агента (защитная функция иммуноглобулина). Классы иммуноглобулиновУ человека известно 5 классов И.: IgM, IgD, IgG, IgE и IgA. Как при дифференцировке В-лимфоцитов, так и в ходе первичного иммунного ответа первыми синтезируются IgM; они эффективно связывают комплемент, служат основными рецепторами антигенов на поверхности зрелых В-лимфоцитов, где присутствуют в виде мономеров; в сыворотке крови находятся в форме пентамеров (включают 5 мономеров, соединённых J-цепями). IgD в следовых количествах присутствуют в сыворотке крови здоровых людей; выполняют функцию мембранных рецепторов на поверхности В- лимфоцитов. IgG доминируют в сыворотке млекопитающих (более 75% от общегоколичества И.) и составляют основу вторичного (развивающегося при повторной встрече с тем же антигеном) иммунного ответа. Они способны проникать через плаценту, обеспечивая иммунную защиту плода. IgG подразделяются на 4 подкласса - IgG1, IgG2, IgG3, IgG4, доля каждого из которых в сыворотке убывает в указанном ряду. Большинство подклассов (IgG1, IgG2, IgG3) способно активировать комплемент. Все подклассы связываются с Fc-рецепторами полиморфно-ядерных лейкоцитов, а IgG1 и IgG3 - также и с Fc-рецепторами моноцитов. IgE взаимодействует с Fc- рецепторами тучных клеток и базофил; при контакте нескольких связанных с рецепторами молекул IgЕ с антигеном эти клетки начинают секретировать серотонин, гистамин и др. медиаторы, что может явиться причиной аллергич. реакции и анафилактич. шока. IgА защищают слизистые оболочки, а также обеспечивают передачу иммунитета от матери к новорождённому, подразделяются на два подкласса: IgА1, присутствующие в осн. в сыворотке крови, и IgА2, содержащиеся в слюне, пищеварит. соке, молозиве, слизистой оболочке носа.Строение молекулы иммуноглобулиновИ. разных классов имеют структурные и функциональные особенности, но общая организация молекул сохраняется у всех И. У большинства изученных видов организмов мономерная молекула И. построена из 4 полипептидных цепей: двух одинаковых тяжёлых (H) и двух одинаковых лёгких (L). Исключение составляют И. сем. верблюдовых и некоторых видов акул, где б. ч. И. содержит только две тяжёлые цепи. Молекулы И. симметричны и представляют собой гликопротеины с чётко выраженной доменной структурой; домены, состоящие примерно из 110 аминокислотных остатков, гомологичны по первичной структуре и имеют характерную укладку цепи (рис. 1). Все 4 цепи соединены дисульфидными связями, которые имеются и внутри доменов, стабилизируя структуру отд. глобулярных областей. N- концевые домены существенно различаются у разных И. и носят назв. вариабельных (V); V H и V L - вариабель ные домены тяжёлой и лёгкой цепей соответственно. Они формируют антигенсвязывающий центр (Fab, или Fv), причём на каждую мономерную молекулу И. приходится по два таких центра. С-концевая часть каждого полипептида в пределах класса изменяется мало и называется константной (С) областью. Она включает один C L -домен у лёгких цепей и три или четыре (в зависимости от класса) С H -домена у тяжёлых цепей. Вариабельность аминокислотных остатков внутри V- доменов наиболее высока в коротких гипервариабельных областях (CDR1-CDR3), разделённых т. н. каркасными участками, на которые приходится ок. 80% всего V- домена. Каркасные участки поддерживают однотипную трёхмерную структуру V- доменов, необходимую для обеспечения контактов гипервариабельных участков с антигенами. Часть молекулы И., состоящую из С-концевых доменов тяжёлых цепей, обозначают как Fc-фрагмент. Он опосредует связывание белков комплемента, взаимодействие с рецепторами лейкоцитов и соединён с остальной частью молекулы И. гибкой «шарнирной» областью, благодаря чему антигенсвязывающие участки И. обладают определённой пространственной свободой. Различают 5 осн. типов тяжёлых цепей (µ, δ, γ, α и ε), которые содержат 450-600 аминокислотных остатков, и 2 типа лёгких (ϰ и λ), включающих ок. 230 аминокислотных остатков. Лёгкие цепи ϰ и λ могут быть соединены с любым типом тяжёлых цепей иммуноглобулина.Гены иммуноглобулиновРазнообразие И. обеспечивается строением их генов и перегруппировками ДНК. В геномах человека и мыши имеются три независимых локуса генов И. - ϰ, λ и H. Каждый из них включает набор генов, кодирующих вариабельные домены и единичные гены константных областей, причём эти группы генов находятся на значит. расстоянии друг от друга. Образование лёгкой и тяжёлой цепей происходит в результате рекомбинации одного из вариабельных генов (V) с геном, кодирующим константный участок (С). Перед каждым геном имеется участок ДНК, кодирующий лидерный пептид (L). Между V- и С-генами находятся последовательности (D-сегменты и J-сегменты), кодирующие небольшие фрагменты полипептидной цепи, входящие в вариабельные области. Огромное разнообразие И. достигается в результате: существования большого числа вариабельных областей V-генов в каждом из H-, ϰ - и λ -локусов; рекомбинации между V-генами, D- и J-cегментами, кодирующими фрагменты вариабельных доменов; разнообразия вариантов, возникающих на границе соединяющихся сегментов как следствие неточного соединения, делеции или включения дополнит. нуклеотидов; комбинаций между лёгкими и тяжёлыми цепями. Дополнит. разнообразие И. определяется соматическими точечными мутациями, происходящими в перегруппированных генах. При созревании В-лимфоцитов имеет место перегруппировка ДНК, приводящая к переносу одного из V-генов к одному из J-сегментов (в случае генов тяжёлых цепей предварительно протекает рекомбинация между D- и J-сегментами, рис. 2). В результате такой V(D)J-рекомбинации происходит сближение вариабельных и константных участков, а также регуляторных элементов транскрипции (промоторов и энхансеров). В ходе развития клеток иммунной системы может происходить изменение класса синтезируемого И. Оно обусловливается генетич. процессом, включающим новую перегруппировку ДНК и носящим назв. «переключение изотипа». При этом происходит замена в зрелом гене тяжёлых цепей одного С H -гена другим. Процесс не затрагивает ни V H -генов, ни генов лёгких цепей, и, следовательно, специфичность И., синтезируемых данной клеткой, остаётся неизменной. В некоторых случаях изменение класса синтезируемой тяжёлой цепи достигается за счёт альтернативного сплайсинга.Применение иммуноглобулиновВ 1975 разработан метод, позволяющий получать клеточные линии (гибридомы), секретирующие гомогенную популяцию моноклональных антител. Последние широко используются для целей диагностики и биотехнологии, но применение для терапии ограничивается тем, что с помощью гибридомной технологии удаётся получать молекулы гл. обр. мышиных И., которые вызывают у человека иммунный ответ. Целью нового направления в области иммунологии, названного «инженерия антител», является создание на основе генов И. не существующих в природе структур с заранее заданными свойствами. Получено неск. типов таких антител. В химерных, или гуманизированных, антителах контактирующие с антигеном участки вариабельных доменов (CDR) взяты от мышиных И., а всё остальное, включая каркасные участки V- доменов, - от И. человека; они обладают низкой иммуногенностью и способны адекватно взаимодействовать с иммунной системой человека. Иммуноадгезины состоят из Fc-фрагмента молекулы И. и лиганда, специфичного к определённому рецептору; такая молекула в результате связывания с рецептором на поверхности инфицированных вирусом клеток и за счёт Fc-фрагмента способна вызывать уничтожение инфициров. клеток лейкоцитами посредством антителозависимой цитотоксичности или действия системы комплемента. Мини-антитела, или Fv- фрагменты, представляют собой лишь часть молекулы И., отвечающую за связывание чужеродного антигена (из-за малого размера они легче проникают в патогенную ткань и обладают пониженной иммуногенностью). Рекомбинантные иммунотоксины и иммуноферменты получают в результате экспрессии генов И., соединённых с генами токсинов или ферментов. И. широко используются в медицине для диагностики, профилактики и лечения разл. заболеваний.Литература Лит.: Деев С.М. Иммуноглобулины. Белки иммунной системы. М., 1997; Ройт А., Бростофф Дж., Мейл Д. Иммунология. М., 2000; Mayer G. Immunoglobulins-structure and function // Microbiology and Immunology on-line / http://pathmicro.med.sc.edu. |
 |