

Л > ЛИ Лист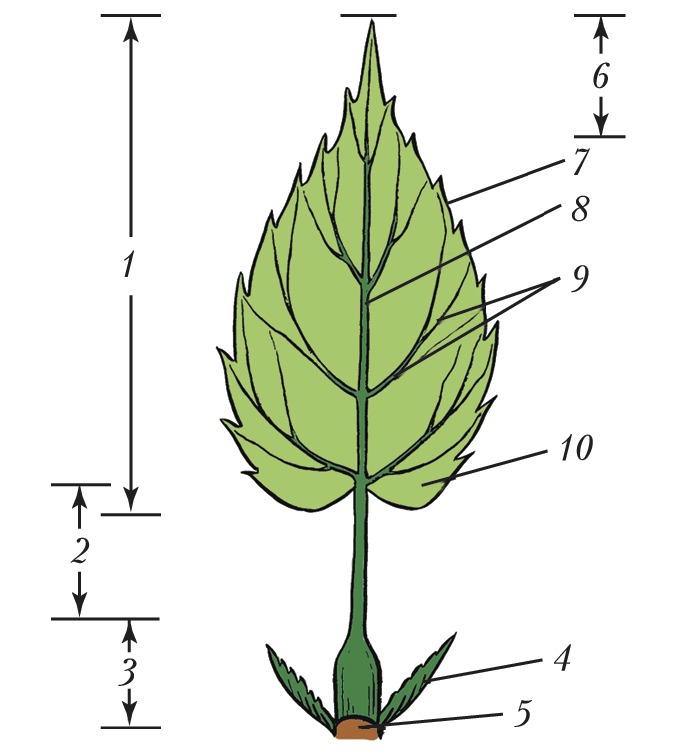  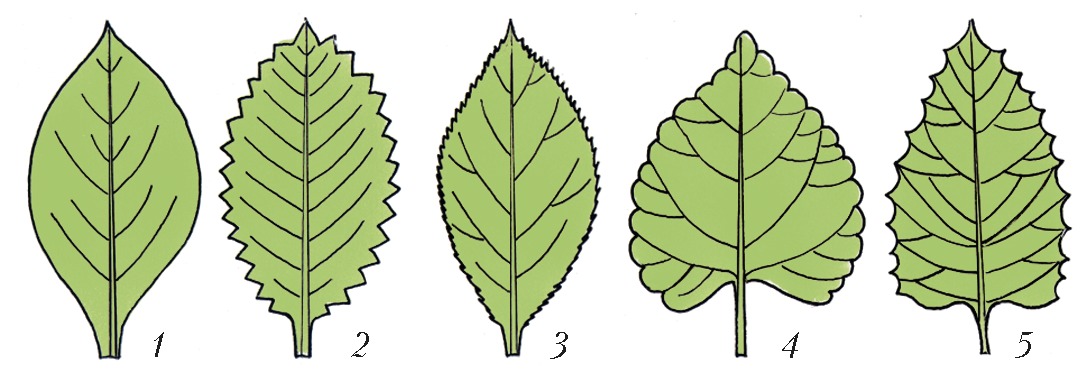 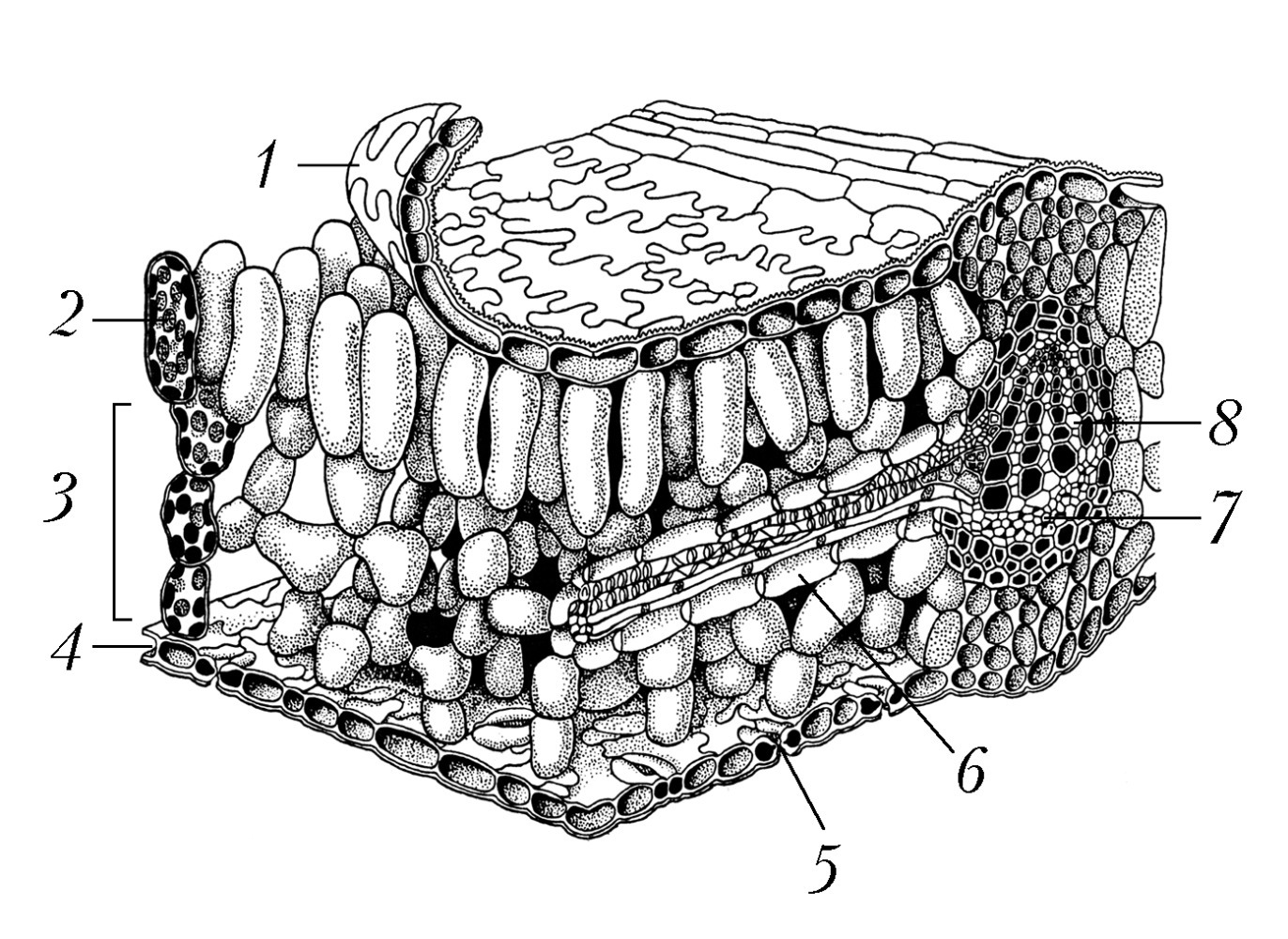  ЛИСТАвторы: А.К. ТимонинЛИСТ, структурный элемент побега растения, выполняющий функции фотосинтеза, транспирации и газообмена. В отличие от стебля и корня, обычно имеет плоскую форму, боковое положение и ограниченный рост. В ходе эволюции Л. возникли как специализир. органы фотосинтеза, в ходе которого осуществляется превращение энергии солнечного света в энергию органич. веществ. Л. сосудистых растений (кроме плауновидных), называемые макрофиллами, образовались из боковых веточек вегетативного тела первичных наземных растений в результате ослабления роста и ветвления веточек, их уплощения и срастания друг с другом краями. Следы такого происхождения сохранились в своеобразном строении Л. папоротников, способных к длительному росту. Л. плауновидных (микрофиллы, или филлоиды) нередко считают разросшимися боковыми выростами, или энациями, на поверхности теломов. У мхов и печёночников Л. (филлидии), по-видимому, сформировались из волосков (трихом). Морфологическое и анатомическое строение листаСр. длина Л. чаще всего колеблется в пределах 3-15 см, однако размеры его варьируют от нескольких мм (у мхов) до 25 м (у некоторых пальм и папоротников). Продолжительность жизни Л., особенно у растений умеренных широт, обычно невелика - ок. 6 мес, хотя у т. н. вечнозелёных растений Л. могут жить дольше (напр., у копытня - 15 мес, у лавра - 3-4 года, у ели европейской - 8-10 лет). В Л., достигших предельного возраста, начинаются процессы старения, ведущие к их отмиранию (см. Листопад). В Л. различают основание, листовую пластинку, прилистники и черешок, которые у разных видов растений развиты в разной степени или отсутствуют (кроме основания) (рис. 1). Основание Л. обычно плохо заметно, но у некоторых видов оно разрастается в трубковидное влагалище, охватывающее стебель (злаки, осоковые, зонтичные), либо остаётся коротким, но сильно утолщается, формируя листовую подушку (ель, пихта и др.). Листовая пластинка - гл. часть Л., как правило, осуществляющая его осн. функции; располагается либо на верхушке основания (сидячие Л. злаков, горечавок и др.), либо на отрастающем от него черешке (черешковые Л. липы, тополя и др.), который, помимо опорной и проводящей функции, может регулировать положение пластинки, изгибаясь по направлению к свету. Развивающиеся по бокам от черешка или пластинки сидячего Л. прилистники обычно остаются мелкими и быстро опадают (яблоня, ива), но у некоторых видов они крупные и напоминают листовую пластинку (горох) или срастаются друг с другом в трубчатый плёнчатый раструб, охватывающий стебель (щавель, гречиха); у мн. видов прилистники вообще не образуются (сложноцветные). У простых Л. одна листовая пластинка, у сложных - две или более, каждую из них называют листочком независимо от размера (рис. 2). Все листочки пальчато-сложного и тройчатосложного Л. располагаются на верхушке черешка, перистосложного Л. - попарно или поочерёдно на рахис е, представляющем собой продолжение черешка. У непарно-перистосложных Л. рахис завершается конечным листочком, у парно- перистосложных Л. такого листочка нет. Бывают также дважды перистосложные и дважды (или многократно) тройчатосложные Л., у которых листочки располагаются на рахисах 2-го и более высоких порядков. По форме, строению основания, верхушки и края Л. (рис. 3), наличию и особенностям строения лопастей листовые пластинки видоспецифичны. В подавляющем большинстве листовая пластинка имеет верхнюю и нижнюю стороны, которые нередко различаются окраской, густотой опушения, рельефом поверхности. У вальковатых Л. листовые пластинки цилиндрические (мн. солянки), а у мечевидных - уплощены с боков и имеют вместо верхней и нижней сторон правую и левую стороны (ирис, гладиолус). Листовая пластинка (рис. 4) с обеих сторон имеет эпидерми с, покрытый кутикулой, а иногда и восковым налётом. Устьица Л., через которые осуществляются газообмен и транспирация, находятся чаще всего только в нижнем эпидермисе (гипостомные Л.), у плавающих по поверхности воды листовых пластинок они расположены в верхнем эпидермисе (эпистомные Л.; кувшинка), а у щирицы, очитка, проса и ряда др. растений - в эпидермисе обеих сторон (амфистомные Л.). Между верхним и нижним эпидермисом находится мезофилл (мякоть Л.), осн. объём которого чаще всего занимает специализир. ткань, обеспечивающая фотосинтез, - хлоренхим а, состоящая из клеток с многочисл. хлоропластами, содержащими зелёный пигмент растений хлорофилл. У мн. листовых суккулентов наряду с хлоренхимой развита водоносная паренхим а, состоящая из клеток, не имеющих хлоропластов. У ряда растений (напр., пшеница, берёза) хлоренхима дифференцирована на палисадную (состоит из 1-3 слоёв клеток, вытянутых перпендикулярно поверхности Л.), которая находится под верхним эпидермисом, и губчатую (с крупными межклетниками), расположенную над нижним эпидермисом. У некоторых тропич. и субтропич. светолюбивых растений палисадная хлоренхима располагается под верхним и над нижним эпидермисами, а губчатая занимает ср. часть мезофилла (эквифациальные Л.: фикус, агава). Соснам, елям и некоторым др. хвойным свойственна складчатая хлоренхима, однотипные клетки которой имеют глубокие впячивания (инвагинации) клеточной мембраны и клеточной стенки. Анатомич. строение листовых пластинок может варьировать у одного и того же растения в зависимости от их освещённости. Напр., у т. н. световых Л., развивающихся в верхних ярусах и на периферии кроны, абсолютная и относит. толщина палисадной хлоренхимы и число слагающих её слоев клеток больше, чем у теневых Л. в глубине кроны. Система проводящих пучков составляет основу жилкования Л. По ксилеме жилок в мезофилл поступают вода и растворённые в ней минер. вещества; по флоэме от Л. оттекают растворы органич. веществ, образовавшихся в процессе фотосинтеза. Чем крупнее пучок, тем более мощная обкладка из паренхимы, а иногда и склеренхимы (механич. ткани), образуется вокруг него. В результате жилка, содержащая такой пучок, выдаётся на нижней стороне листовой пластинки в виде валика. Обкладки пучков крупных жилок достигают верхнего и нижнего эпидермиса и разделяют хлоренхиму листовой пластинки на отд. участки. Через черешок и основание Л. проводящие пучки (листовые следы) входят в стебель и соединяются с его проводящей системой. У ряда растений с особым С4-типом фотосинтеза (просо, щирица) обнаружены особые структуры вокруг мелких жилок, т. н. кранц-структур ы, представляющие собой обкладку пучка из крупных клеток с гигантскими хлоропластами, к которой венцом примыкают удлинённые клетки хлоренхимы. Форма, размеры и анатомич. строение Л. обычно отражают приспособленность к определённым экологич. условиям.Функциональное приспособление листаУ мн. видов растений Л. одного побега могут сильно различаться строением и функцией. Низовые чешуевидные Л., состоящие только из листового основания, защищают конус нарастания молодого побега с развивающимися на нём срединными Л., которые впоследствии и осуществляют фотосинтез. Верховые Л. побега (имеют в пазухах цветки или соцветия) обычно мелкие чешуевидные, но иногда крупные, яркоокрашенные, участвующие в привлечении насекомых-опылителей к цветкам (иван- да-марья, пуансеттия). Срединные Л. чаще всего однотипные, но есть виды, у которых срединные Л. на побеге резко различаются (гетерофиллия, анизофиллия). У ряда видов (ясень и др.) сильно отличаются срединные Л. молодых и взрослых растений. У некоторых растений все или только часть Л. видоизменены в связи с расширением или сменой ими функций. Напр., Л., преобразованные в плотные, жёсткие почечные чешуи, защищают покоящиеся почки в неблагоприятные для роста сезоны. Л., видоизменённые в колючки (у крыжовника, кактусов), предохраняют растения от поедания травоядными животными. С помощью Л., преобразованных в усики, цепляющиеся лианы закрепляют свои побеги на др. растениях (горох). Толстые мясистые зелёные Л. листовых суккулентов (агава, алоэ) наряду с фотосинтезом запасают воду, а мясистые чешуи луковиц (лук, лилия, тюльпан), не содержащие хлорофилла и не фотосинтезирующие, - углеводы. Щитовидные Л. дали начало мешковидным Л., которые насекомоядные растения (саррацения, гелиамфора) используют для ловли и переваривания насекомых, а эпифитная дисхидия Рафлса - для сбора и хранения дождевой воды. В эволюции мн. высших растений спорангии (органы, в которых образуются споры) переместились на Л., и возникли спорофиллы. У одних видов (плаун-баранец, щитовник) последние сходны с обычными Л., а у других - резко от них отличаются (страусник, гинкго и др.).Формирование листьев в онтогенезе и листорасположениеПервые листовые органы семенных растений - семядоли зародыша. В онтогенезе Л. формируются из экзогенных боковых выростов на конусе нарастания побега. Л. папоротников присущ длительный верхушечный рост; у семенных растений верхушечный рост прекращается очень рано, и листовые зачатки в почке растут гл. обр. за счёт краевой меристемы; при этом создаётся в миниатюре будущая форма пластинок. Л. появляются в основании конуса нарастания побега через определённые промежутки времени (пластохрон ы) поодиночке, парами напротив друг друга или группами по 3-4 (редко больше) вокруг стебля на равных расстояниях друг от друга. Соответственно различают очередное (рассеянное), супротивное и мутовчатое листорасположение. Тип листорасположения (филлотаксис а) обычно постоянен и специфичен для вида. Очередные Л. (у дуба, берёзы, злаков, зонтичных) закладываются и располагаются по т. н. осн. генетич. спирали (воображаемой линии), соединяющей последовательно возникающие Л. При этом каждый Л. образуется на максимально возможном для определённого вида растений угловом расстоянии от предшествующего листа, что уменьшает затенение Л. друг друга. Тем не менее некоторые Л. оказываются строго друг над другом, и через них можно мысленно провести прямую (ортостих у), параллельную стеблю. На укороченных побегах с очередными Л. можно мысленно провести контактные парастихи - линии, соединяющие тесно сближенные Л., не соседствующие друг с другом на основной генетич. спирали. На каждом побеге удаётся построить по несколько право- и левозакрученных контактных парастих в строго определённом числовом соотношении. При супротивном (у клёна, сирени, губоцветных) и мутовчатом (у олеандра, элодеи) листорасположении число возможных ортостих вдвое превышает число Л. на одном узле стебля, причём Л. соседних узлов оказываются на соседних ортостихах, т. е. строго против промежутков между Л. соседних узлов, и потому минимально затеняют друг друга. Взаимное затенение Л. ещё более уменьшается благодаря их разному размеру, скручиванию междоузлий побега и искривлению черешков. В результате проекция Л. одного побега способствует макс. использованию света; она подобна флорентийской мозаике, отчего её и называют листовой мозаикой (рис. 5).Использование листьевЛ. находят разнообразное применение в хозяйств. деятельности человека. Их употребляют в пищу (салат, щавель, укроп, лук, петрушка и др.), используют для выкармливания гусениц шелкопряда и вместе с др. частями растений - на корм скоту. Из Л. мн. растений получают лекарственные вещества; они входят в состав лекарственных сборов. Л. герани, мяты и др. растений - источник эфирных масел, используемых в парфюмерной и кондитерской пром-сти. Из Л. некоторых растений добывают дубящие вещества. Л. являются сырьём в чайной и табачной пром-сти. В тропиках из Л. мн. растений получают грубое волокно, Л. пальм используют как кровельный материал. См. также ст. Фотосинтез.Литература Лит.: Федоров А. А., Кирпичников М. Э., Артюшенко З.Т. Атлас по описательной морфологии высших растений. Лист. М.; Л., 1956; Гамалей Ю. В., Куликов Г.В. Развитие хлоренхимы листа. Л., 1978; Васильев Б.Р. Строение листа древесных растений различных климатических зон. Л., 1988; Гамалей Ю.В. Транспортная зависимость эволюции листа двудольных // Журнал общей биологии. 2004. Т. 65. № 5. |
 |